|
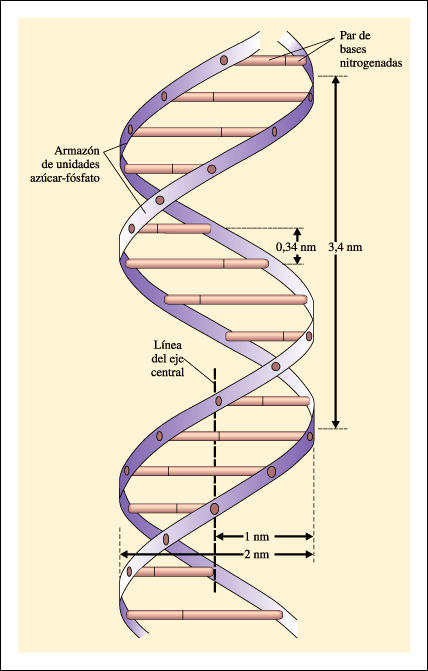
La estructura de doble hélice del DNA, como fue presentada en 1953 por Watson y Crick. |
|
|
ADN : El modelo de Watson y Crick |
James Watson y Francis Crick se dedicaron a examinar y constrastar todos los datos existentes acerca del ADN, y a unificarlos en una síntesis significativa.
En el momento en que Watson y Crick comenzaron sus estudios, ya había un cúmulo de abundante información.e sabía que la molécula de DNA era muy grande, también muy larga y delgada, y que estaba compuesta de nucleótidos que contenían las bases nitrogenadas adenina, guanina, timina y citosina.
Los físicos Maurice Wilkins y Rosalind Franklin habían aplicado la técnica de difracción de rayos X al estudio del DNA. Las fotografías obtenidas mostraban patrones que casi con certeza reflejaban los giros de una hélice gigante.
También fueron cruciales los datos que indicaban que, dentro del error experimental, la cantidad de adenina (A) es igual que la de timina (T) y que la de guanina (G) es igual que la de citosina (C): A=T y G=C.
A partir de estos datos, algunos de ellos contradictorios, Watson y Crick intentaron construir un modelo de DNA que concordara con los hechos conocidos y explicara su papel biológico. Para llevar la gran cantidad de información genética, las moléculas debían ser heterogéneas y variadas.
Reuniendo los diferentes datos, los dos científicos fueron capaces de deducir que el DNA es una doble hélice, entrelazada y sumamente larga.
Si se tomase una escalera y se la torciera para formar una hélice, manteniendo los peldaños perpendiculares, se tendría un modelo grosero de la molécula de DNA. Los dos parantes o lados de la escalera están constituidos por moléculas de azúcar y fosfato alternadas. Los peldaños perpendiculares de la escalera están formados por las bases nitrogenadas adenina, timina, guanina y citosina. Cada peldaño está formado por dos bases, y cada base está unida covalentemente a una unidad azúcar-fosfato. En la doble hélice, las bases enfrentadas se aparean y permanecen unidas por puentes de hidrógeno, esos puentes relativamente débiles que Pauling había encontrado en sus estudios sobre la estructura de las proteínas. De acuerdo con las mediciones efectuadas mediante rayos X, las bases apareadas (los peldaños de la escalera) debían ser siempre combinaciones de una purina con una pirimidina.
Cuando Watson y Crick analizaron los datos, armaron modelos reales de las moléculas usando alambre y hojalata, ensayando dónde podía encajar cada pieza en el rompecabezas tridimensional. A medida que trabajaban con los modelos, advirtieron que los nucleótidos situados en cualquiera de las cadenas de la doble hélice podían acoplarse en cualquier orden o secuencia. Dado que una molécula de DNA puede tener miles de nucleótidos de largo, es posible obtener una gran variedad de secuencias de bases diferentes, y la variedad es uno de los requisitos primarios del material genético.
Notaron también que la cadena tiene dirección: cada grupo fosfato está unido a un azúcar en la posición 5' -el quinto carbono en el anillo de azúcar- y al otro azúcar en la posición 3' -el tercer carbono en el anillo de azúcar-. Así, la cadena tiene un extremo 5' y un extremo 3'.
Sin embargo, el descubrimiento más excitante ocurrió cuando Watson y Crick comenzaron a construir la cadena complementaria. Encontraron otra restricción interesante e importante. No solamente las purinas no podrían aparearse con purinas, ni las pirimidinas con pirimidinas, sino que, a causa de las estructuras particulares de las bases, la adenina sólo podía aparearse con la timina, formando dos puentes de hidrógeno (A=T) y la guanina solamente con la citosina, formando tres puentes de hidrógeno (G=C). Las bases apareadas eran complementarias.
Las dos cadenas corren en direcciones opuestas, es decir, la dirección desde el extremo 5' al 3' de cada cadena es opuesta y se dice que las cadenas son antiparalelas. Aunque los nucleótidos dispuestos a lo largo de una cadena de la doble hélice pueden presentarse en cualquier orden, su secuencia determina el orden de los nucleótidos en la otra cadena. Esto es necesariamente así, porque las bases son complementarias (G con C y A con T).
|
La estructura de doble hélice del DNA, como fue presentada en 1953 por Watson y Crick. |
|
El armazón de la hélice está compuesto por las unidades azúcar-fosfato de los nucleótidos. Los peldaños están formados por las cuatro bases nitrogenadas, adenina y guanina (purinas) -cada una de ellas con un anillo doble en su estructura- y timina y citosina (pirimidinas), más pequeñas, cada una con un sólo anillo. Cada peldaño está formado por dos bases. El conocimiento de las distancias entre los átomos fue crucial para establecer la estructura de la molécula de DNA. Las distancias fueron determinadas con fotografías de difracción de rayos X del DNA, tomadas por Rosalind Franklin y Maurice Wilkins.
|
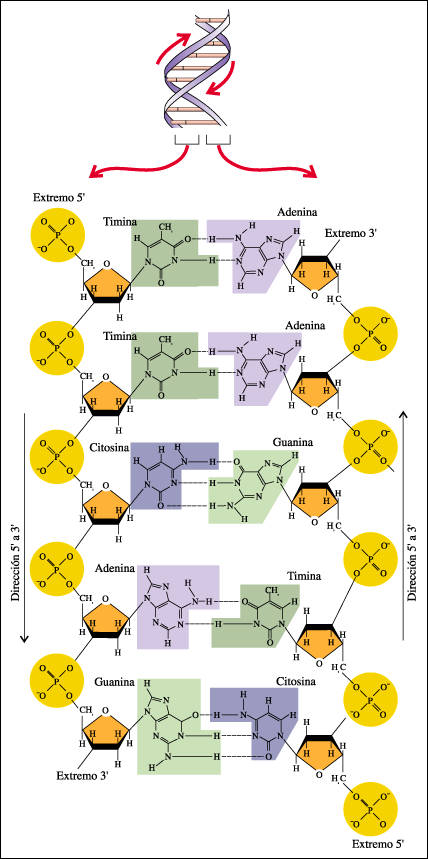
La estructura de doble cadena de una porción de una molécula de DNA. |
|
|
En el modelo de Watson y Crick, cada nucleótido consiste en un azúcar desoxirribosa, un grupo fosfato y una base púrica o pirimídica. Nótese la secuencia repetida azúcar-fosfato-azúcar-fosfato que forma el esqueleto de la molécula. Cada grupo fosfato está unido al carbono 5' de una subunidad de azúcar y al carbono 3' de la subunidad de azúcar del nucleótido contiguo. Así, la cadena de DNA tiene un extremo 5' y un extremo 3' determinados por estos carbonos 5' y 3'. La secuencia de bases varía de una molécula de DNA a otra.Las cadenas se mantienen unidas por puentes de hidrógeno (representados aquí por guiones) entre las bases. Nótese que la adenina y la timina pueden formar dos puentes de hidrógeno, mientras que la guanina y la citosina pueden formar tres. Dados estos requerimientos de enlace, la adenina puede aparearse sólo con la timina y la guanina sólo con la citosina. Así, el orden de las bases en una cadena -TTCAG- determina el orden de las bases en la otra cadena -AAGTC. Las cadenas son antiparalelas, es decir, la dirección desde el extremo 5' a 3' de una es opuesta a la de la otra.
Fuente : Invitación a la Biología, Curtis